
| 일 | 월 | 화 | 수 | 목 | 금 | 토 |
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| 15 | 16 | 17 | 18 | 19 | 20 | 21 |
| 22 | 23 | 24 | 25 | 26 | 27 | 28 |
- BacterPhage
- β-Mannanases
- Non-starch polysaccharide
- CTCBIO
- Low Protein
- Disease challenge
- Xylannase
- β-mannan
- Charoen Pokphand Group
- CTCZYME
- Nutrition
- Diseases of Swine
- Danish Crown
- bacteriophage
- CP Group
- LAYER
- C.P. Group
- Digestibility
- egg quality
- split feeding
- Palm kernel expeller
- Gut health
- calcium availability
- Pig
- 김인배박사
- Exolution
- broiler
- β-mannanase
- Phytase
- Beta-mannanases
- Today
- Total
CTCBIO Animal Bioscience Blog
Effects of exogenous phytase and β-mannanase on ileal and total tract digestibility of energy and nutrient in palm kernel expeller-containing diets fed to growing pigs 본문
Effects of exogenous phytase and β-mannanase on ileal and total tract digestibility of energy and nutrient in palm kernel expeller-containing diets fed to growing pigs
Kwon, KiBeom 2023. 3. 27. 11:35
Abstract
본 연구의 목적은 10% palm kernel expellers 함유한 사료를 급여한 돼지에서 에너지와 영양소의 명백한 회장 및 총관 소화율에 대한 phytase 및 β-mannanase의 효과를 결정하는 것이다.
기본 식단은 주로 옥수수, 대두박, 무기 P가 없는 야자박을 기반으로 했습니다.
0 또는 1000 U/kg의 phytase 보충 처리구 2개와 0 또는 1600 U/kg의 β-mannanase 처리구 2개로 2 × 2 요인 처리 배열로 4가지 실험 식단을 준비했다.
개시 체중 70.9kg(SD = 11.9) 돼지 8마리에 회장 T-캐뉼라를 장착하고 4회 식이 요법과 4주기로 복제된 4 × 4 라틴 스퀘어 디자인으로 식이 실험을 진행했다.
각 기간에서 d7에 분변을 수집하고 d8 및 d9에 회장 소화물 샘플을 수집했습니다.
Phytase의 보충은 P의 ileal apparent digestibilities 계수(CIAD)와 총 total tract apparent digestibilities (CTTAD) 를 각각 0.161과 0.191 증가시켰습니다(P < 0.01).
β-mannanase 첨가는 건물, 유기물 및 에너지의 CIAD를 0.035 ~ 0.038(P < 0.05), 에너지의 CTTAD를 0.019(P < 0.05) 증가시켰습니다.
그러나 영양소의 소화율에서는 phytase와 β-mannanase 사이의 상호작용이 관찰되지 않았다.
Phytase는 aspartic acid, glutamic acid, serine의 CIAD를 0.021 ~ 0.029(P<0.05) 증가시켰고 β-mannanase는 leucine과 phenylalanine의 CIAD를 각각 0.018과 0.019 증가시켰다(P<0.05).
결론적으로 phytase와 β-mannanase는 각각 인과 유기 영양소의 소화율을 향상시켰고, 2개의 효소는 아미노산 소화율을 약간 향상시켰다.
1. Introduction
돼지 사료의 phytase 보충은 사료내 phytic acids를 가수분해하여 P의 소화율을 향상시킵니다(Mroz et al., 1994; Sands and Kay, 2007; Agudelo Trujillo et al., 2010).
돼지 사료내 mannan은 영양소 이용을 방해하는 것으로 제안되었습니다(Rainbird et al., 1984).
옥수수-대두박(SBM) 기반 식이에 대한 β-mannanase 보충이 영양소 소화율 및 성장 성능에 미치는 영향이 연구되었습니다(Pettey et al., 2002).
흥미롭게도 palm kernel expellers (PKE; 244g/kg)의 mannan 함량은 옥수수(9g/kg), 보리(27g/kg), 밀(20g/kg), SBM(20g/kg), 유채박(30g/kg), 면실박(15g/kg) 보다 훨씬 많다.
Palm kernel expellers와 palm kernel meal은 사료 비용을 줄이기 위해 돼지사료에 사용되어 왔다(Kim et al., 2001).
그러나 PKE 함유 돼지 사료에 대한 β-mannanase의 효과는 아직 보고되지 않았다.
또한, PKE 함유 사료를 먹인 돼지의 영양소 소화에 대한 phytase와 β-mannanase의 상호 작용 효과에 대한 정보는 제한적입니다.
따라서 본 연구의 목적은 phytase 와 β-mannanase 또는 두 가지 효소로 옥수수-SBM-PKE 기반 사료를 보충하는 것이 돼지 성장에 의한 에너지 및 영양소의 명백한 회장 및 총 소화관 소화율에 미치는 영향을 테스트하는 것이다.
2. Materials and methods
본 실험의 프로토콜은 건국대학교 동물실험실 관리위원회(KU11082)의 검토 및 승인을 받았다.
2.1. Animals and experimental design
개시 체중이 70.9kg(SD = 11.9)인 교잡 수퇘지 8마리를 사료 첨가제로 사용되는 phytase와 β-mannanase의 효과를 측정하기 위해 사용하였다.
공시가축은 4개의 다이어트와 사각형당 4개의 주기로 복제된 4×4 라틴 사각형 디자인에 할당되었다(Kim and Kim, 2010).
돼지는 Stein 등(1998)에서 채택한 절차를 사용하여 T-cannula in the distal ileum 외과적으로 장착했다.
시험돈군은 급이기와 니플 급수기가 장착된 돈방에 개별적으로 수용했다..
2.2. Diets and ingredients
0 또는 1000 U/kg의 보조 대장균 유래 phytase 처리구 2개와 0 또는 1600 U/kg의 β-만나나아제 처리구 2개로 2×2 요인 처리 배열로 4개의 실험 식단을 준비했다.
Beta-mannanase는 Luria 국물에서 자란 Bacillus subtilis에서 생산되었습니다.
기본 식단은 67% 옥수수, 20% SBM 및 10% PKE를 기반으로 했습니다.
Phytase, β-mannanase 또는 두 효소 제품 모두 옥수수 전분을 희생하여 기본 식단에 추가되었습니다(Table 1).

2.3. Sample collection
각 기간은 9일 동안 진행하였다.
각 기간의 초기 적응 기간은 6일간 이었다.
d7일에 대변을 수집하고 d8 및 9일에 7시간 동안 회장 소화물 샘플을 수집했습니다.
Ilealdigesta 수집은 Kim et al(2009)의 절차 기반으로 했습니다.
유선 비닐 백(Whirl-Pak®)을 캐뉼라 배럴에 부착하고 백으로 흐르는 다이제스타를 수집했습니다.
소화액이 채워질 때마다 또는 적어도 30분에 한 번씩 백을 제거했습니다.
수집된 샘플은 소화관에서 박테리아에 의한 아미노산(AA) 분해를 방지하기 위해 -18℃에 보관되었습니다.
2.4. Chemical analysis
만난 농도에 대해 총 9개의 사료 성분을 분석하였다.
성분 샘플은 60℃에서 건조하였다.
샘플은 72%(w/w) H2SO4를 사용하여 1시간 동안 가수분해되었습니다.
그런 다음 샘플을 H2SO4 농도가 1 N이 되도록 증류수로 희석하고 121°C에서 45분 동안 배양했습니다.
가수분해물의 만난 함량은 증발 광산란 검출기와 Shodex 설탕 칼럼 SP0810(8.0 mm × 300 mm; Showa Denko K.K., Tokyo, Japan)을 사용하여 결정되었습니다.
실험 종료 시, 동결된 회장 소화관과 대변을 동결 건조하고 화학적 분석 전에 1mm 스크린(Cyclotec 1093; Foss Tecator AB, Hoganas, Sweden)을 통해 미세하게 분쇄했습니다.
Kim et al(2009)이 기술한 폭탄 열량계(Model C2000, IKA® , Germany)를 사용하여 식이, 회장 소화관 및 대변 샘플을 총 에너지에 대해 분석했습니다.
모든 샘플은 AOAC(2005)에 기술된 바와 같이 건물(DM; method 930.15), 유기물(OM; method 942.05), P(method 964.06) 및 Cr(method 990.08)에 대해 분석되었습니다.
식이 및 회장 소화물 샘플의 아미노산 농도도 AOAC(2005, method 982.30)에 설명된 방법을 사용하여 분석했습니다.
Methionine은 가수분해 전 밤새 cold performic acid oxidation 후 methionine sulfone 으로 분석되었습니다.
Tryptophan은 110ºC에서 22시간 동안 4N NaOH 가수분해 후 분석되었습니다.
2.5. Calculations and statistical analysis
실험 식이에서 에너지에 대한 회장 겉보기 소화율(CIAD) 계수와 총관 겉보기 소화율 계수(CTTAD)는 Stein 등(2007)에서 채택한 다음 방정식에 따라 계산되었습니다.
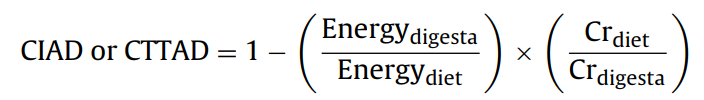
여기서 CIAD 및 CTTAD는 각각 에너지의 CIAD 및 CTTAD를 나타냅니다.
Energydigesta는 회장 또는 대변 소화물의 GE이고, Energydiet는 식이의 에너지 농도이고, Crdiet는 식이의 Cr 농도이며, Crdigesta는 회장 또는 대변 소화물의 Cr 농도입니다.
DM, OM 및 P의 CIAD 및 CTTAD도 동일한 방정식을 사용하여 계산되었습니다.
에너지의 단위는 MJ/kg으로 표시하였고, 영양소 농도는 g/kg으로 표시하였다.
Coefficient of hindgut digestibility 는 CTTTD에서 CIAD를 빼서 에너지와 영양소를 계산했습니다.
실험 데이터는 SAS(SAS Institute Inc., Cary, NC, USA)의 GLM 절차를 사용하여 분석하였다.
모델에는 phytase, β-mannanase 및 phytase와 β-mannanase 사이의 상호 작용이 포함되었습니다.
초기 모델에는 복제, 복제 내 동물, 복제 내 기간도 무작위 변수로 포함되었으나, 이러한 무작위 변수는 반응 기준에 대한 중요성이 없기 때문에 최종 모델에서 제외되었습니다.
돼지는 실험 단위였으며 유의성을 결정하기 위해 alpha level 0.05를 사용했습니다.
3. Results and discussion
2기간에 1마리를 제외한 모든 동물은 건강하였고, 제공된 사료를 쉽게 섭취하였으며, 실험 기간 동안 평균 체중이 10.4kg 증가하였다.
Phytase 보충은 P의 CIAD 및 CTTAD를 각각 0.161 및 0.191 증가시켰습니다(P < 0.01; Table 2).

Beta-mannanase는 DM, OM 및 에너지의 CIAD를 0.035–0.038(P < 0.05), DM, OM 및 에너지의 CTTAD를 0.019–0.026(P < 0.01) 증가시켰습니다.
DM, OM, 에너지, P(P>0.10)의 CIAD와 CTTAD에서 phytase와 β-mannanase 사이의 상호작용은 관찰되지 않았다.
소화기 말단 소화율은 phytase 또는 β-mannanase에 의해 영향을 받지 않았다(P > 0.20).
Phytase 보충은 인 소화율 개선은 많은 이전 보고서와 일치합니다(Sands and Kay, 2007; Agudelo Trujillo et al., 2010; Kerr et al., 2010).
옥수수, SBM 및 PKE는 각각 총 P kg당 808, 535 및 712g의 피테이트 결합 P를 함유하고 있으며(NRC, 2012), 이는 피타아제 기질이 식단에 풍부함을 나타냅니다.
Phytase 보충 없는 사료에서 P의 CTTAD는 0.325로 옥수수, SBM 및 PKE에서 P의 CTTAD가 각각 0.26, 0.39 및 0.39임을 고려할 때 합리적인 범위 내에 있습니다(NRC, 2012).
Pettey 등(2002)은 DM과 에너지 소화율에 차이가 없다고 보고했지만, 옥수수-SBM 기반 식이와 β-mannanase 첨가 기본 식이 간에는 차이가 없었습니다.
Beta-mannanase 보충에 의한 DM, OM 및 에너지 소화율의 현저한 개선이 본 실험에서 관찰되었습니다.
이러한 불일치는 본 연구에서 사용된 식이에서 β-mannanase에 대한 기질의 양으로 설명될 수 있습니다.
PKE의 만난 함량은 옥수수, 밀, 보리, SBM, 카놀라박 또는 면실박보다 8배 이상 높았습니다.
본 연구에서 β-mannanase의 효과는 아마도 사료내 β-mannanase에 대한 기질이 풍부하기 때문에 감지되었습니다.
기질에 대한 β-mannanase의 작용을 명확히 하기 위해 섬유 또는 만난 소화율에 대한 추가 분석이 필요합니다.
Kiarie et al(2010)의 실험에서 보조 xylanase와 β-glucanase는 phytase와 상호 작용하여 에너지의 회장 소화율을 증가시켰습니다.
이는 기질의 양으로 설명할 수 있습니다.
Kiarie et al(2010)의 연구에서 보리는 β-glucan과 phytate를 모두 함유하고 있으며 밀은 arabinoxylan과 phytate를 모두 충분히 함유하고 있다.
따라서 carbohydrase와 phytase는 단일 성분에서 기질을 상승적으로 가수분해할 수 있습니다.
그러나 본 연구에서 옥수수와 SBM은 실험 식이에서 전체 phytate 결합 P kg당 555g과 299g의 phytate 결합 P를 제공한 반면 PKE는 다량의 만난을 함유하고 있으며, NRC(2012)의 각 성분에 대한 포함률 및 피테이트 결합 P 농도를 기반으로 사료에서 총 피테이트 결합 P kg당 146g의 피테이트 결합 P를 제공했습니다.
따라서 phytase와 β-mannanase의 상호작용을 관찰하지 못한 이유는 본 실험에서 두 효소의 주요 기질 성분이 다르기 때문일 수 있다.
PKE를 더 많이 포함하거나 phytate와 mannan을 다량으로 포함하는 성분에 대한 추가 연구를 통해 phytase와 β-mannanase 사이의 가능한 상호 작용을 밝힐 수 있습니다.
추가 phytase는 Asp, Glu 및 Ser의 CIAD를 0.021에서 0.029까지 증가시켰고(P < 0.05; 표 3), β-mannanase는 Leu 및 Phe의 CIAD를 각각 0.018 및 0.019 증가시켰습니다(P < 0.05).

His, Trp, Tyr에 대한 겉보기 회장 소화율 계수는 phytase가 없을 때 β-mannanase에 의해 감소한 반면 소화율 값은 phytase가 있을 때 β-mannanase에 의해 증가하여 phytase × β-mannanase 상호 작용을 초래했습니다(P < 0.05 ).
AA 소화율에 대한 피타아제의 효과는 문헌에서 일관성이 없습니다(Adeola and Sands, 2003; Kiarie et al., 2010; Morales et al., 2012).
Phytase가 현재 실험에서 일부 AA 활용을 개선했지만 개선 정도는 실질적으로 중요하지 않을 수 있습니다.
마찬가지로, AA 활용은 이 연구에서 β-mannanase에 의해 최소한의 영향을 받는 것으로 보입니다.
Kim 등(2006)은 β-galactosidase, β-mannanase 및 β-mannosidase로 구성된 보충 carbohydrase complex에 의한 AA의 CIAD 개선을 보고했습니다.
Yin et al(2001)은 또한 보충 탄수화물이 다양한 성분 기반 식단에서 AA의 CIAD를 개선했다고 기록했습니다.
그러나 두 연구 모두에서 보충 탄수화물에 의한 AA 소화율의 개선은 대부분 5% 미만이었으며, 이는 현재 결과와 일치합니다.
방향족 AA의 CIAD에 대한 phytase와 β-mannanase의 상호작용 효과는 흥미롭습니다.
그러나 AA의 CIAD에 대한 β-mannanase의 효과가 추가 피타아제에 의해 어떻게 강화되는지는 알 수 없습니다.
결론적으로, phytase와 β-mannanase는 2개의 보충 효소 사이에 주요한 상승적 상호작용 없이 각각 야자핵 추출물 함유 식이의 P와 유기 영양소 소화율을 향상시켰습니다.



